A few years ago, I was asked to explain lactic acid fermentation in sourdough, and the difference between homo- and heterofermentation. Not an easy task, partly because I wasn't satisfied that I knew enough, or that I could reconcile what I was reading in bread-baking books with what I had learned in school. To sort it out, I had to dig deeper into the scientific literature. Answers are there in bits and pieces, although not in a context that is easy to make sense of. As I plugged away at deciphering current microbiology textbooks and scientific research, I started to see things in a new light. And so now, I want to share what I've learned with those who wish to know more.
First, I'd like to introduce the concept of a metabolic pathway. On paper, a metabolic pathway can be illustrated in a flow diagram that represents a sequence of enzyme-controlled chemical transformations. While the pathways in this discussion start with sugar and finish as various end-products, there are several intermediate compounds formed along the way as one thing is converted to the next. The names may be intimidating at first glance, but don't let them scare you. Knowing their chemical reactions and what all the compounds are is not as important here as understanding their overall purpose, which is to produce energy for the organism. Like all living things, microbes need energy to perform the tasks that enable them to live, grow and multiply.
Some pathways generate more energy than others. Through respiration, glucose and oxygen are turned into carbon dioxide and water via the Krebs cycle, also known as the tricarboxylic acid or TCA cycle. You may have seen it before if you've studied biology, because it's the same pathway we humans use. It is aerobic, meaning that oxygen (O2) is involved, and it generates far more energy than any fermentation pathway. Whenever oxygen is available, respiration is favored by facultative anaerobes like yeasts, because they will always take the path that generates the most energy under the prevailing conditions. For the most part though, bread dough is anaerobic (without oxygen), and fermentation is an alternative pathway that doesn't require oxygen. When yeasts ferment sugars, they produce alcohol (ethanol) in addition to carbon dioxide. Fermentation produces much less energy than respiration, but it allows microorganisms to carry on when no oxygen is available, or they lack the ability to respire as is the case with lactobacilli.
Bacterial fermentation is more varied than fermentation by yeast. Bacteria produce organic acids that contribute, for good and bad, to the quality of bread. Controlling acid balance and degree of sourness is something that artisan bakers strive to do, so it may be useful to understand where the acids come from and how their production can be influenced by things that are within the baker's control. In yeasted breads, acids come in small doses from naturally occurring bacteria present in flour and commercial yeast. (Fresh yeast generally has more bacterial inhabitants than dried, and whole grain flours more than refined.) In sourdough breads, acid-producing bacteria are supplied in much greater numbers from starter. There are many different species and strains of bacteria found in various types of starters, and because they produce lactic acid while fermenting sugar, they fall under the heading of Lactic Acid Bacteria (LAB).
Lactic acid bacteria common to sourdoughs include members of Leuconostoc, Pediococcus, Weissella and other genera. But by far, the most prevalent species belong to the very large and diverse genus, Lactobacillus. Based upon how they ferment sugars, lactic acid bacteria can be sorted into three categories. Please bear with me now, because while these terms may look impossibly long and technical, they are actually self-descriptive. Take homofermentative LAB for example. Homo-, meaning "all the same," refers to the end product of fermentation (by lactic acid bacteria), which is only, or "all" lactic acid. Heterofermentative then, means "different" or mixed end products. As lactic acid bacteria, heterofermentative LAB produce lactic acid, but they also produce carbon dioxide gas, alcohol or acetic acid as well.
SUGAR STRUCTURE
As carbo-hydrates, sugars are made up of carbon (C) and water, which is composed of hydrogen (H) and oxygen (O). The hydrogen and oxygen atoms are arranged in various configurations around a chain of carbon atoms which form the structural backbone of the molecule. The carbon chain may be various lengths, but sugars common in bread fermentations are of the 5- and 6-carbon types, referred to generically as pentoses and hexoses, respectively. Glucose and fructose are examples of hexoses. Pentoses are sugars such as arabinose and xylose.
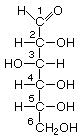
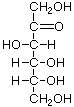
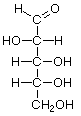
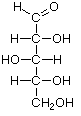
Glucose Fructose Arabinose Xylose
Pentoses and hexoses can exist in the chain form, or in a ring structure which forms when dissolved in water. Single sugars, or monosaccharides, are often linked together into larger carbohydrates of two or more units. Disaccharides, containing two sugars, are important in bread fermentations. Maltose, which is made up of two glucose molecules, is the free-form sugar most abundant in dough. Sucrose, another disaccharide consists of one glucose and one fructose.
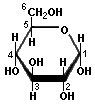
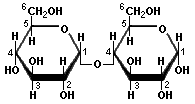
Glucose Maltose
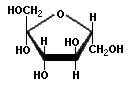
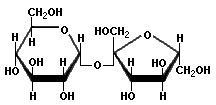
Fructose Sucrose
-Sugars illustrated by Antonio Zamora. For a more complete explanation, with diagrams of starches and pentosans, please see his lesson, "Carbohydrates - Chemical Structure" at: http://www.scientificpsychic.com/fitness/carbohydrates.html
Sugars that can be fermented, and their end-products are variable from one species of LAB to the next. But the key lies in the structure of the sugar---particularly, the number of carbon atoms in the backbone of the molecule. Homofermentative LAB can only ferment 6-carbon sugars. In the homofermentative pathway, a hexose is processed and split into two identical 3-carbon pieces, which are passed down through the reaction sequence and transformed into lactic acid molecules. In contrast, heterolactic fermentation is based on 5-carbon sugars. Pentoses may be used directly, although more often, a hexose is cut down by removing one of its carbons. The extra carbon is cast off in the form of carbon dioxide gas, and the remaining 5-carbon molecule is split unequally into 3- and 2-carbon units. The 3-carbon piece is turned into lactic acid, while the 2-carbon piece will become either ethanol or acetic acid. Up to this point, heterolactic fermentation doesn't produce as much energy as homolactic, but it does give an advantage over homofermentative LAB, which cannot utilize pentose sugars.
Additional energy can be produced by turning the 2-carbon piece into acetic acid, but it requires the assistance of another substance. The term for this is co-metabolism, meaning that two substrates are used simultaneously---a hexose for its carbon backbone, and a co-substrate to facilitate the formation of acetic acid and generation of additional energy. The co-substrate can be one of a number of things including oxygen, citrate, malate, short chain aldehydes, oxidized glutathione, fructose and 5-carbon sugars. In the absence of co-substrates, the 2-carbon piece is turned to ethanol instead. Alternatively, when pentose sugars are fermented (used as the carbon source), acetic acid may be produced without the help of co-substrates.
Some lactobacilli can use oxygen as a co-substrate. Some cannot, and are inhibited by aerobic conditions. In any case, there is a small amount of oxygen in dough only at the beginning of fermentation, and generally not enough to affect acetic acid production to any extent. Likewise, citrate and malate aren't naturally present in significant amounts, and pentose utilization varies by species and strain as well as availability. While all these things may be used to the extent that they are present, it turns out that fructose is generally the one most available in bread dough.
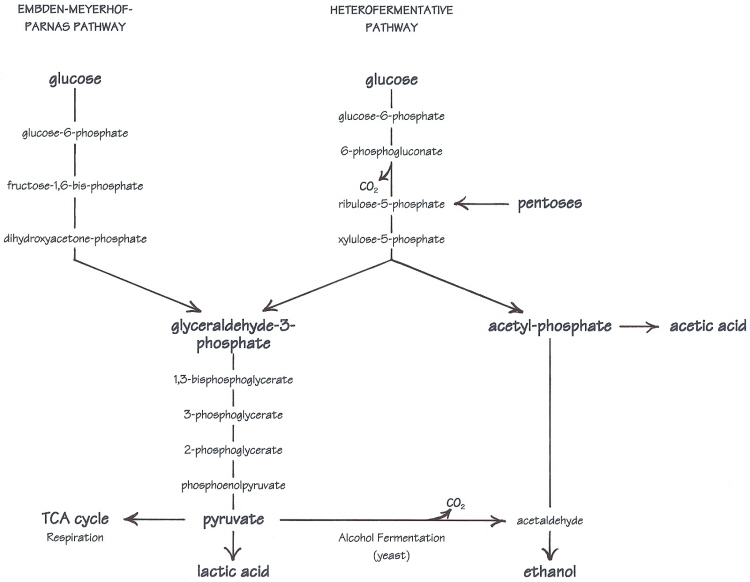
All of the pathways in this discussion are glycolytic pathways. Glycolysis is the conversion of glucose to pyruvate, which is the springboard to both respiration and alcohol fermentation in yeast, to lactic acid fermentation in LAB, and to many biosynthetic pathways (manufacture of compounds used in other life processes). Oxygen is not required, so glycolysis is especially important to microorganisms that ferment sugars, like the yeast and bacteria which grow in the anaerobic environment of sourdough.
Homofermentative lactobacilli share the same glycolytic pathway with yeasts---the Embden-Meyerhof-Parnas, or EMP pathway. But in contrast to alcohol fermentation, pyruvate is reduced to lactic acid. In either of the two pathways here, the sugars are split into smaller molecules---two identical 3-carbon units (glyceraldehyde-3-phosphate) in the EMP pathway, or a 3- and a 2-carbon unit in the heterofermentative pathway. The 3-carbon pieces all follow the same path to become pyruvate and then lactic acid, while the 2-carbon acetyl-phosphate on the other side of the heterofermentative pathway can become either ethanol or acetic acid.
Glucose is not the only sugar that can be utilized. With appropriate enzyme systems, other sugars can be converted into glucose or one of the intermediates in the pathway such as glucose-6-phosphate (or in the case of pentose sugars, ribulose-5-phosphate). The ability to use other sugars varies by species and strain. Most sourdough lactic acid bacteria ferment glucose preferentially, but Lactobacillus sanfranciscensis separates maltose into a glucose-1-phosphate and a glucose. The glucose-1-phosphate portion is converted to glucose-6-phosphate to enter the heterofermentative pathway, and glucose is excreted from the cell.
In addition to obligately hetero- and homofermentive, there is a third type of lactobacilli characterized as facultatively heterofermentive. These are lactobacilli that are not restricted to one pathway or the other, but can use both. Facultative heterofermenters switch back and forth between the homo- and heterofermentative pathways depending upon which sugars are available. In general, they ferment hexoses via the homofermentive route, and pentoses heterofermentively. Most will use the hexose sugars first, although some strains ferment pentoses preferentially. Many co-metabolize fructose with maltose through the heterofermentative pathway, but use the homofermentative pathway when only maltose is available.
To put all this technical information to practical use, we need to consider factors that influence LAB activity and pathway selection. The end products are determined by the species and available sugars, which for lean doughs, depend upon the flour and the activity of enzymes. Whole grain and high extraction flours can affect acidification in two ways. First, the higher mineral (ash) content serves as a natural buffer system, which allows bacteria to produce more acid before the pH drops low enough to slow their growth. And second, grains supply pentose sugars in the form of pentosans. Although rye flours are best-known for these, pentosans are also present in wheat and other grains. (But, because they occur in the outer layers of the kernel, they are largely removed along with enzymes and many other substances in the milling of refined flours.) Cereal enzymes act on pentosans to some degree, freeing pentose sugars like xylose and arabinose that heterofermenters may be able to use according to species and strain. Pentoses will increase acetic acid production if they can be fermented or co-metabolized, either one.
Acidification is also influenced by hydration and temperature. Contrary to popular belief, all three groups of sourdough lactobacilli prefer wetter doughs a bit on the warm side, many growing fastest at about 90ºF or a little higher. For the homofermentive species producing only lactic acid, increasing activity by raising the hydration and/or temperature will increase acid production. Decreasing activity by reducing hydration or by retarding will slow production. There is a direct relationship between activity and lactic acid. During heterofermentation, for each molecule of glucose consumed, one lactic acid is produced, along with one carbon dioxide (if a hexose is fermented), and either one ethanol or one acetic acid. But under wetter, warmer conditions, where sugars are metabolized more rapidly, the tendency is toward lactic acid and alcohol production in obligate heterofermenters, and all lactic acid (homofermentation) in the facultative heterofermenters. Lactic acid production is directly related to activity during heterofermentation just as in homofermentation, even if only half the rate.
At lower hydrations and temperatures (lower activity), more acetic acid is produced, but not because of temperature per se. Acetic acid production is influenced indirectly by temperature, in that it affects the kinds of sugars available. The fructose that drives acetic acid production, is liberated from fructose-containing substances in flour, largely through the enzyme activity of yeast. And, because lower temperatures are more suited to yeast growth than higher, more fructose is made available to the bacteria at lower temperatures. At the same time, the bacteria are growing and using maltose more slowly, so the demand for co-substrates goes down as the fructose supply goes up. The ratio of acetic acid to ethanol and lactic acid goes up, because a higher percentage of the maltose is being co-metabolized with fructose. Reducing hydration has a similar effect of slowing the bacteria more than yeast, which I believe is the real basis for increased acetic acid production in lean breads made with refined flours.
Contrary to myth, the species that grow in sourdough starters are not tied to geographic location, but rather to the traditional practices in the different regions. Several organisms go into the mix, but the environment created inside the starter from the combination of flour, temperature and maintenance routines is what determines which ones will thrive. In type I, or traditional sourdoughs (i.e., those maintained by continuous refreshment at room temperature), the obligately heterofermentive Lactobacillus sanfranciscensis is the species most frequently and consistently found---not just in San Francisco where it was first discovered, but all around the world. And so it deserves special attention.
Lactobacillus sanfranciscensis is fairly unique among the obligately heterofermentive lactobacilli, in that it ferments no pentose sugars. And unusual among lactic acid bacteria in general, because it prefers maltose over glucose. But it will co-metabolize fructose with maltose to produce acetic acid. L. sanfranciscensis converts maltose into one glucose-6-phosphate which enters the heterofermentative pathway, and a glucose which is excreted back into its surroundings. This is a good arrangement for common sourdough yeasts, since maltose is the most abundant sugar in wheat doughs, and some lack the ability to break it down for themselves. Yeasts and other bacteria that can ferment maltose, generally prefer glucose. And so by providing glucose to competing organisms, L. sanfranciscensis actually helps to conserve the maltose for itself---just one of the ways in which it gets along well with other sourdough microorganisms, and perhaps one of the reasons it is found so often.
Alternate pathways are a recurring theme in the microbial world, because microorganisms have less ability to control their environment or to leave when conditions become difficult. They sometimes have to switch gears to survive. In that effort, lactic acid bacteria will utilize whichever fermentation pathway that generates the most energy within their capabilities and resources. In order of preference, the hierarchy seems to be heterofermentation with co-substrates (forming lactic acid and acetic acid), followed by homofermentation (all lactic acid) and heterofermentation without co-substrates (lactic acid and ethanol).
While traditional sourdough starters usually support one or more strains of Lactobacillus sanfranciscensis, it is often found in combination with the facultatively heterofermentive Lactobacillus plantarum, many strains of which can either ferment or co-metabolize at least one pentose sugar. Various other obligate and facultatively heterolactic acid bacteria are also common (obligately homofermentive LAB are only transient in the startup process and do not persist in established type I starters). Sourdough starters are sensitive ecosystems with complex associations of lactic acid bacteria, and combinations can be highly variable from one starter to the next. Lactic acid fermentation is as complex and varied as the organisms involved, and so sourdough processes may need to be optimized on a starter by starter basis.
- Debra Wink
Bibliography
Arendt, Elke K., Liam A.M. Ryan, and Fabio Dal Bello. 2007. Impact of sourdough on the texture of bread. Food Microbiology 24: 165-174.
De Vuyst, Luc and Marc Vancanneyt. 2007. Biodiversity and identification of sourdough lactic acid bacteria. Food Microbiology 24:120-127.
Doyle, Michael P., Larry R. Beuchat, and Thomas J. Montville. 2001. Microbial physiology and metabolism, p. 19-22; Lactose metabolism, p. 653-655. Food Microbiology Fundamentals and Frontiers, 2nd ed. American Society for Microbiology Press, Washington, DC.
Gänzle, Michael G., Michaela Ehmann, and Walter P. Hammes. 1998. Modeling of growth of Lactobacillus sanfranciscensis and Candida milleri in response to process parameters of sourdough fermentation. Applied and Environmental Microbiology 64:2616-2623.
Gänzle, Michael G., Nicoline Vermeulen, Rudi F. Vogel. 2007. Carbohydrate, peptide and lipid metabolism of lactic acid bacteria in sourdough. Food Microbiology 24:128-138.
Gobbetti, M., P. Lavermicocca, F. Minervini, M. De Angelis, and A. Corsetti. 2000. Arabinose fermentation by Lactobacillus plantarum in sourdough with added pentosans and "alpha-L-arabinofuranosidase: a tool to increase the production of acetic acid. Journal of Applied Microbiology 88:317-324.
Holt, John G., Noel R. Krieg, Peter H. A. Sneath, James T. Staley, and Stanley T. Williams. 2000. Regular, nonsporing gram-positive rods, p. 566. Bergey's Manual of Determinative Bacteriology, 9th ed. Lippincott Williams & Wilkins, Philadelphia, PA.
Katina, Kati. 2005. Sourdough: a tool for the improved flavour, texture and shelf-life of wheat bread. VTT Technical Research Centre of Finland.
Ng, Henry. 1972. Factors affecting organic acid production by sourdough (San Francisco) bacteria. Applied Microbiology 23:1153-1159.
Paramithiotis, Spiros, Aggeliki Sofou, Effie Tsakalidou, and George Kalantzopoulos. 2007. Flour carbohydrate catabolism and metabolite production by sourdough lactic acid bacteria. World J Microbiol Biotechnol 23:1417-1423.
Wing, Daniel, and Alan Scott. 1999. Baker's Resource: Sourdough Microbiology, p. 230. The Bread Builders. Chelsea Green Publishing Company, White River Junction, VT.
This article was first published in Bread Lines, a publication of The Bread Bakers Guild of America. Vol. 15, Issue 4, Dec. 2007.
Revised: November 4, 2009

























 Blueberries and cream cheese wrapped in a sweet yeasted dough. Yes, it really is as good as it sounds, and it is making me hungry again just sitting here thinking about it.
Blueberries and cream cheese wrapped in a sweet yeasted dough. Yes, it really is as good as it sounds, and it is making me hungry again just sitting here thinking about it. 












